

1. 서 론
2. 자료와 방법
3. 결 과
4. 토 의
4.1 용존 규소의 농도와 동해 컨베어 벨트
4.2 일본분지 저층에서의 일정한 용존 규소 연직분포
4.3 일차생산에 대한 용존규소 제한 가설
4.4 동해에서 용존 규소의 생지화학적 순환이 지닌 함의
1. 서 론
규소는 지구의 지각에서 산소 다음으로 풍부한 원소로서 암석의 주성분이다. 하지만 해수에서 규소는 열역학적 평형(포화농도, 약 1000μmol kg-1)에 크게 못 미치는 불포화 상태(< 200μmol kg-1)에 있다. 그 결과로 용존 규소는 해수의 주성분이 아니다. 해수에서 용존 규소의 주된 화학종은 약산인 규산(Si(OH4) 또는 H4SiO4; pKa ~9)이고 해수의 정상적인 pH 범위에서 약 5%만 해리하여 음이온인 H3SiO4-로 존재한다(Sarmiento and Gruber, 2006).
암석을 이루는 용존 규소광물(aluminosilicates)과는 달리 생규소(biogenic silica)는 비정질 광물로서 열역학적으로 매우 불안정하지만 대표적인 규질 생물인 규조가 살아있는 동안에는 유기물 피복으로 보호되어 용해되지 않으며 사후에 미생물에 의한 유기물 피복이 분해되기 시작하면서 용해가 진행된다(Bidle and Azam, 1999). 해수가 용존 규소에 대해 크게 불포화 상태이므로 생규소(SiO2·nH2O)는 표면 혼합층(상부 약 100 m) 안에서 생산량의 절반이 넘게 규산(Si(OH)4)으로 빠르게 용해되어 재순환된다(Trégure and De La Rocha, 2013).
수중에서 생규소의 분해 속도는 생규소 자체의 속성(질량 대비 표면적, 수화도, 불순물 함유도)과 해수 환경(수온, 염분, 압력, pH)과 포화도에 지배된다(Schmidt et al., 2001). 용해를 피해 심해로 침강한 생규소는 수온이 낮아지고, 불포화도와 실효 표면적이 줄면서 분해가 매우 느려져서 대부분은 퇴적 후에 표층 퇴적층에서 분해되어 상층 해수로 공급되는 것으로 알려졌다(Trégure et al., 1995). 하지만 생규소의 용해 속도는 실험실 실험으로 예측한 값의 약 1/5 수준으로 해양은 규소에 대해 크게 불포화된 상태로 유지되고 있다. 생규소의 퇴적물 매장 효율은 전 지구적으로 약 3%로 추정된다(Ragueneau et al., 2000).
강물을 비롯한 다양한 경로로 해양으로 공급되는 규산과 쉽게 규산으로 용해되는 생규소의 플럭스(~9.4 Tmol Si yr-1)는 규질 생물의 생규소 생산(240 Tmol Si yr-1)과 심해로의 송출 플럭스(~105 Tmol Si yr-1)에 비해 매우 작아서 용승 플럭스(~99.3 Tmol Si yr-1)에 의해 규질 생물(규조, 규질편모류, 해면과 방산충)의 생산력이 유지되고 있다(Trégure and De La Rocha, 2013). 또한 생규소 생산은 퇴적물로 최종 제거되는 플럭스(6.3 Tmol Si yr-1)의 대략 40배에 이르는 규모로서 해양의 규소 순환에서 생물 개입의 중요성을 단적으로 보여주고 있다. 따라서 용존 규소의 해양에서의 연직분포는 생규소의 생물 생산과 용해에 의해 주도된다.
지구 역사에서 규소의 순환은 신생대 에오세부터 용존 규소를 사용하는 규조류가 번성하면서 현재와 같은 모습을 갖추게 된 것으로 이해되고 있으며 규소는 현대 해양에서 질소와 인과 함께 기초생산에 대량으로 요구되는 영양염 가운데 하나이다. 해양에서 용존 규소는 규조(diatom)의 일차생산력을 조절하고 다른 한편으로는 이들이 만들어 낸 생규소가 부력 조절을 통해 해양의 생물학적 탄소펌프에 절반 가량 기여하는 탄소 침강 플럭스를 만들어내기 때문에(Nelson et al., 1995) 탄소 순환과 밀접하게 연계되어 있다.
규산의 재활용도는 매우 높아서 최종적으로 퇴적물로 매장되기 전에 약 25회 생물에 의해 동화되는 것으로 추정된다(Trégure and De La Rocha, 2013). 규산의 해양 체류시간은 약 10,000년 규모로서 질소보다는 길고 인보다는 짧다. 생규소의 생산량 대비 매장 효율은 약 3%로서 탄소의 약 0.1%에 비해 30배에 달해(Ragueneau et al., 2000) 생물학적 탄소펌프를 연구하는 지화학자들의 주목을 받게 되었다. 그 결과로 지난 세기 중반부터 해양에서 규소의 생지화학적 순환에 대한 이해가 크게 증진되었다(Tréguer and De La Rocha, 2013 and references there in).
생규소의 매장 효율은 해역에 따라 크게 차이가 나서 생규소 생산력이 높은 해역에서는 오히려 매장 효율이 낮게 나타나는 생산과 보존이 상반되어 나타나는 ‘규소 역설’이 제기된 바 있다. 이는 생규소의 용해 속도에 관한 연구의 중요성을 지적한다. 생규소의 용해는 자체 요인인 용해 메커니즘과 심해 송출과 같은 외부적인 해양 현상의 복합적인 결과이다.
표층 해양에서 전형적인 용존 규소의 농도는 10μmol kg-1 미만이고 심해의 평균 농도는 약 73μmol kg-1 수준인데 우리나라 근방의 경우에는 북태평양의 심해수는 평균보다 높은 170μmol kg-1, 동해 심해수는 83 μmol kg-1, 동중국해는 130 μmol kg-1를 보인다(Isshiki et al., 1991). 연구 지역인 동해는 “축소판 대양”이라는 별칭에 어울리게 자체적으로 심해수를 만들어 순환시키는 컨베어 벨트를 가지고 있다. 하지만 해수가 드나드는 해협은 모두 수심이 얕아서(< 200 m) 북서태평양과 심해수의 교환은 사실상 단절되어 있고 큰 강이 직접 연결되어 있지 않다. 그러므로 동해에서 물질의 생지화학적 순환은 대체로 독자적일 수밖에 없어 보인다. 규소의 관점에서는 동해는 닫힌 계에 매우 가까운 특성을 지닌 자연이 제공한 천혜의 실험실이기도 하다. 이런 맥락에서 매장 효율이 높은 것으로 알려진 규소가 동해에서 어떻게 분포하고 표층으로 원활하게 되돌려지는지 여부를 알아보는 것은 앞으로 기후변화로 유발될 해수순환의 바뀜과 생태계에 대한 영향과 그리고 생지화학적 순환에 관련하여 의미가 있다고 판단된다.
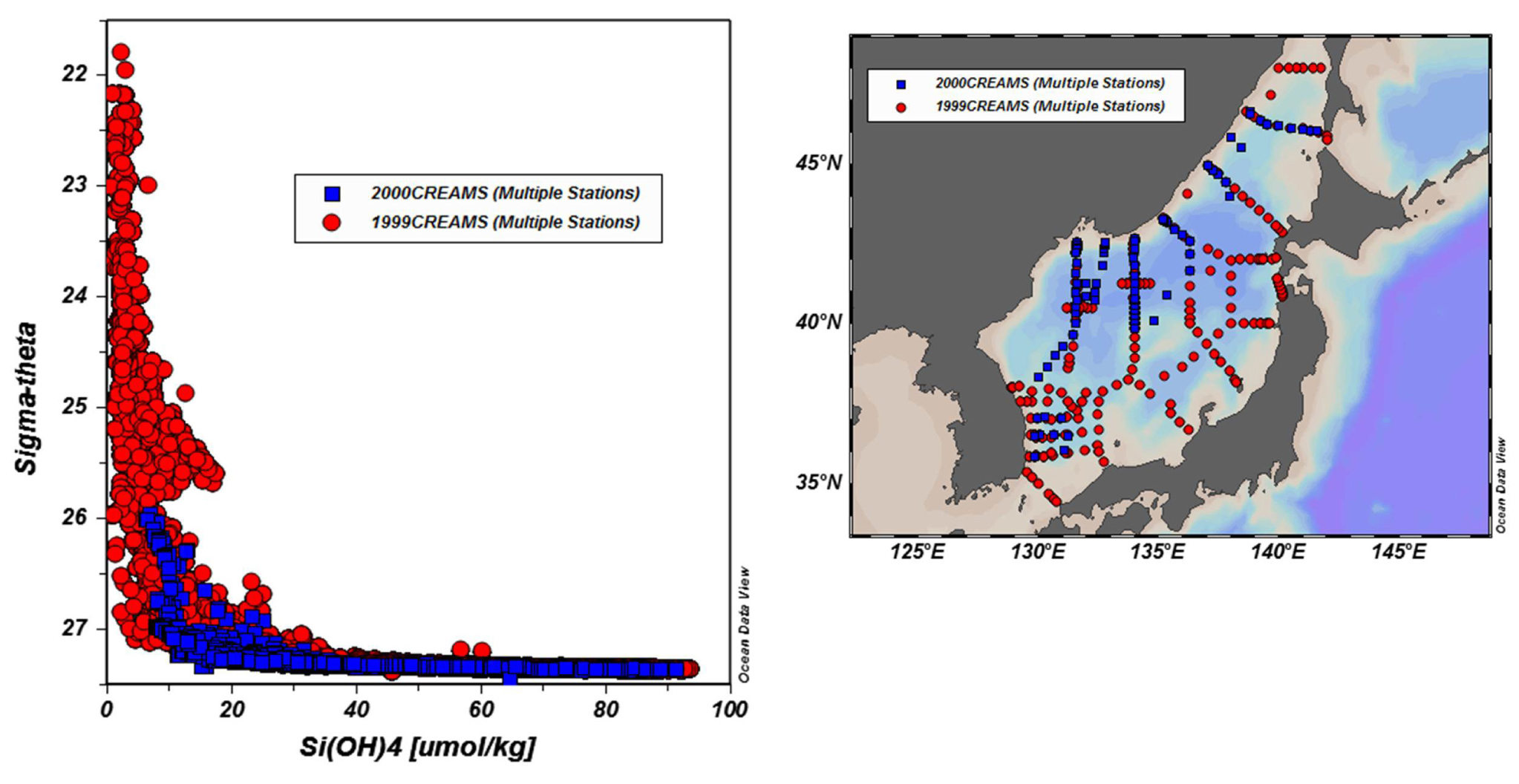
지금까지 동해의 용존 규소 연직분포를 다룬 논문은 희소하다. Kido and Nishimura(1973)는 동해가 지닌 거의 닫힌 계인 특성에 주목하여 동해의 자료로서 해양에서의 규소의 재생을 일반화하려고 시도한 바 있다. 이 논문은 CTD-rosette 채수 시스템이 도입되기 전인 1970년도 조사 자료를 사용했다. Kim et al.(2010)은 1999년도 ONR-JES 조사 자료를 가지고 인산염 대 용존 규소 비의 연직분포에서 밀도약층 하부에 불연속면이 존재함을 보고하였다. 두 논문에서는 20세기 후반부터 축적된 생규소의 용해 거동에 대한 연구결과가 반영되지 않았다.
이 논문에서는 Kido and Nishimura(1973)의 1970년도 용존 규소 연직분포 자료와 30년 후의 ONR-JES 관측 자료를 최근에 알려진 동해 해수순환의 특성에 맞추어 해석해 보았다. 또한 용존 규소의 연직분포가 질산염과 인산염과는 어떻게 다른지를 용해와 해양순환의 관점에서 고찰하였다. 그리고 전 대양과 동해에서의 규소 순환을 대조하여 동해에서의 규산 분포의 특성이 생지화학적 순환에 대해 지닌 함의를 고찰하여 향후 연구 주제를 발굴하는데 초점을 맞추었다.
2. 자료와 방법
용존 규소을 비롯한 영양염은 1999-2000년도에 수행된 ONR-JES 프로그램의 공개된 현장 조사 자료를 사용하였다. ONR-JES 프로그램은 현대적인 관측 기법으로 동해를 가장 광역적으로 조사한 자료를 제공하였으므로 이를 기준으로 이전 자료를 비교 고찰하였다. SBE 911 CTD-rosette 시스템으로 수온, 염분, 용존산소가 측정되고 해수가 채취되었다. ONR-JES 영양염 자료는 스크립스 해양연구소의 ODF 소속 분석자가 분절 흐름식 영양염 자동분석기로 측정하였다. 영양염 자료에 오류표지가 붙어 있거나 규산과 인산염 자료 가운데 누락된 것이 있는 자료는 분석에서 제외시켰다. Kido and Nishimura (1973)에 쓰인 용존 규소 자료는 자체 개발한 방법으로 측정한 값이다. 사용된 영양염은 측정 방법에서 차이가 있지만 측정값이 정확하다고 가정하고 사용하였다.
3. 결 과
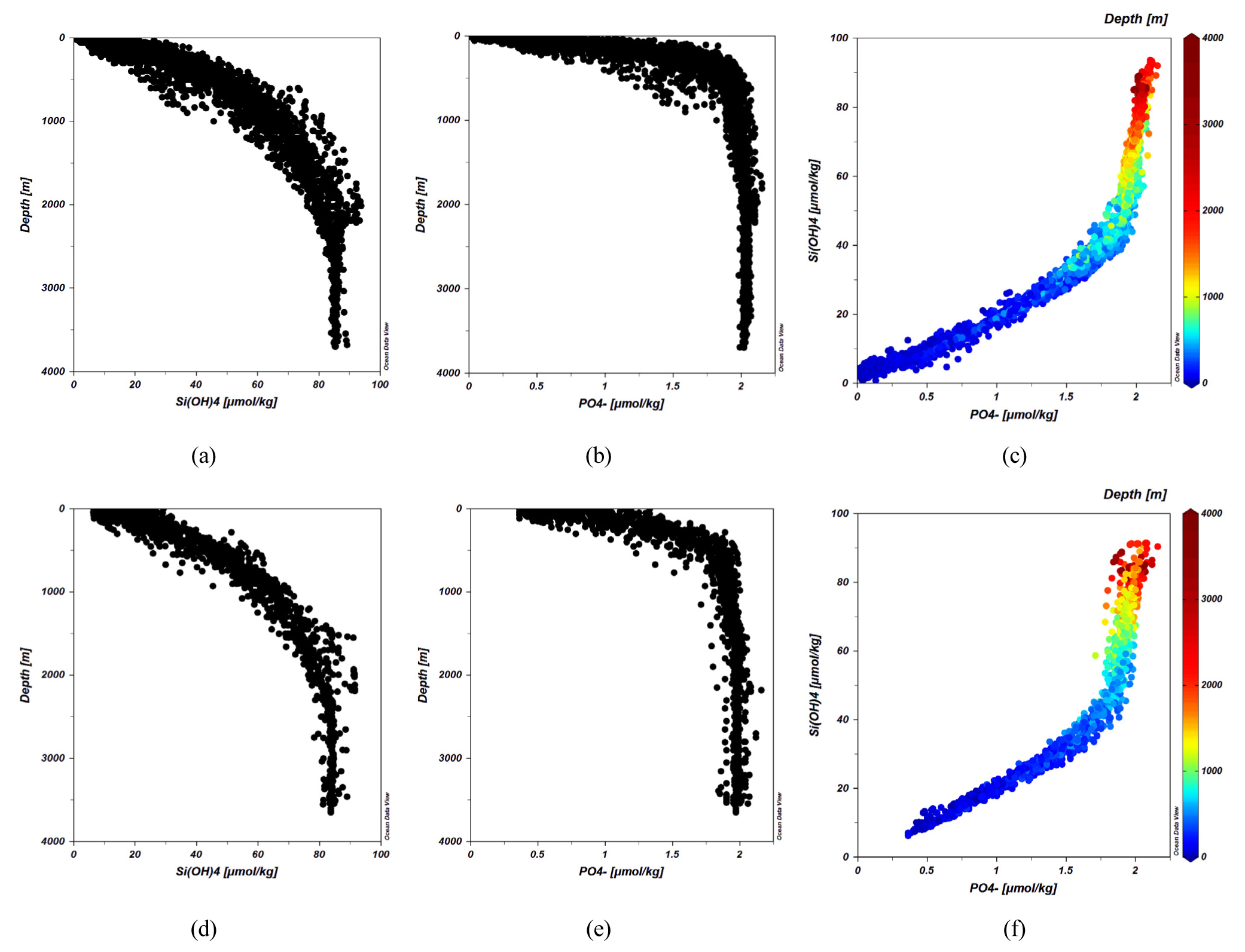
ONR-JES 1999년도 여름철 관측과 2000년도 겨울철 관측 자료를 보면 용존 규소의 농도는 표층 해수에서 낮고 약 2300 m 수심까지 계속 증가하다 그 보다 깊은 곳에서는 증가 추세가 멈추고는 조금 감소하는 추세로 바뀌는데 분산이 매우 적은 일정한 값을 보인다(Fig. 1(a) and (d)). 동해에서 수심이 2300 m를 넘는 곳은 일본분지에 주로 존재하므로 이는 일본분지에 국한된 용존 규소의 분포 특성으로 해석되어야 한다. 일본분지를 예외로 하면 용존 규소의 농도는 바닥까지 계속해서 증가한다고 볼 수 있다. 일본분지가 예외적인 연직분포를 보이는 것에 대해서는 토의에서 다루었다. 용존 규소 농도의 최대값의 최빈값은 대략 85 μmol kg-1로서 북태평양의 2000 m에서 3,000 m 사이의 농도(~120 μmol kg-1)보다는 훨씬 낮으며 이는 둘 사이의 심층수의 교환이 단절된 것을 반영한다.
용존 규소와 달리 수주에서 침강하는 유기물의 산화로 생무기화되는 인산염은 상층에서 급격히 증가하다가 1000 m보다 얕은 수심에서 증가 추세가 멈추고 상층에 비해 값이 거의 일정하며 분산이 줄어든 분포를 보이다가 2300 m보다 깊은 곳에서는 용존 규소과 마찬가지로 약간 감소하는 추세를 보인다(Fig. 1(b) and (e)). 용존 규소의 최대농도 깊이가 인산염의 것보다 깊은 이유는 미생물이 효소를 이용해 유기물을 분해해서 재생되는 인산염이 단순히 화학적 포화도에 지배되어 용해되는 생규소 껍데기보다 빠른 속도로 분해되기 때문으로 해석된다.
두 영양염의 연직분포 특성의 차이는 인산염 대 용존 규소 그래프에서 더욱 뚜렷하게 나타난다(Fig. 1(c) and (f)). 인산염 대 용존 규소의 곡선은 두 개의 기울기가 확연하게 다른 선형 추세선이 혼합 구간에서 부드러운 곡선으로 연결되는 것으로 보여지는데 Kido and Nishimura(1973)의 논문에서는 이를 하나의 지수함수적 곡선으로 해석하였다. 이전 해석이 몇 개 정점의 자료에 근거한 것에 대비해서 ONR-JES 자료의 총 수가 100배 이상 많으므로 경향을 더 제대로 보여주는 것으로 해석할 수도 있고 경과한 기간 사이에 심층순환의 변동에 기인할 수도 있다. 최근 수십년 동안 축적된 용존산소의 자료를 보면 심층수에서 농도가 일방적으로 줄고있고 용존산소 최소층의 깊이가 깊어지고 있어서(Gamo et al., 2014) 해수 교체(ventilation) 양상의 변화가 주도했을 가능성이 높다.
인산염 대 용존 규소 추세선의 기울기는 상층수에서는 겨울철 혼합과 여름철 성층으로 인해 계절적 차이가 나타나지만 대략 1:20 부근이고, 심층수에서는 크게 도약해서 1:100 가까이로 나타난다. 이는 상층(< 300 m)에서는 인산염과 용존 규소가 함께 증가하는데 반해 심층(1000 m ~2300 m)에서는 인산염의 농도 증가는 정체되고 용존 규소만 증가하는 양상을 보이는 것을 반영한다. 두 선형 추세선은 대략 중층(수심 300 m에서 1000 m 구간)에서 용승과 확산에 따른 혼합의 결과로 비선형 전이 구간이 만들어지는 것으로 판단된다. 한편 수심이 2300 m를 넘는 일본분지 저층의 자료는 인산염과 용존 규소의 수심에 따른 농도 변화가 크게 줄어들어서 인산염대 용존 규소의 비 자료는 거의 하나의 점에 수렴해서 심층수 추세선의 전반적인 기울기에는 기여하지 않는다. 울릉분지와 야마토분지의 심층에서는 일본분지 저층에서 보이는 용존 규소가 일정한 저층 구간이 발달해있지 않다.
인산염대 용존 규소 비의 두 추세선이 교차하는 수심은 분지에 따라 500 m에서 800 m 구간이다. 이 수심은 일본분지에서 가장 깊고 울릉분지와 야마토 분지에서는 얕아지는데 분지의 기후 특성의 차이에서 비롯된 것으로 보인다. 아한대 수역인 일본분지에서는 아열대 수역인 울릉분지와 야마토분지보다 겨울철 냉각에 의한 혼합이 더 깊은 곳까지 일어나는 것이 원인이었을 것으로 여겨진다. 한편 2010년 이후 EAST-I 관측 자료는 분지별 차이가 크게 줄어서 500 m 부근인데(미발표자료) 일본분지의 심층수 형성이 약화된 결과로 해석된다.
생규소의 절반 이상이 분해되는 것으로 알려진 표면혼합층 아래에서 용존 규소 농도의 연직 분포는 수온과 밀도성층 구조가 합세하여 주도하는 것으로 해석된다. 표면 혼합층을 빠져나와 침강하는 생규소 입자는 수온이 상대적으로 높고 침강 속도가 느린 밀도약층의 상부에서 주로 분해가 일어나며 그 아래에서는 수온이 낮고 입자의 침강속도가 빨라져서 수중에서 분해가 훨씬 더디게 일어나는 것으로 보인다. 심층수에서 인산염의 농도가 증가하는 추세는 거의 멈추는데 비해 규소의 농도는 40μmol kg-1에서 80μmol kg-1 사이로 계속 증가한다. 이는 심층에서 유기물의 분해와 바닥 퇴적물에서 재생된 인산염의 유입이 함께 미약함을 알려준다. 그에 비해 용존 규소는 퇴적 표면에서 용해된 용존 규소가 심층수로 공급되어 나오므로 일본분지를 제외하고는 바닥까지 계속해서 증가한다. 이로부터 심층에서 인산염의 농도가 균일하게 나타나는 것이 심층 해수의 원활한 혼합의 결과가 아니라는 것을 알려준다. 만일 심층 혼합의 결과라면 용존 규소 또한 마찬가지로 일정한 값을 가져야 하는데 그렇지 않기 때문이다. 저층에서 일본분지의 예외적인 분포는 토의에서 다루었다.
인산염 대 용존 규소 비의 분포를 하나의 매끄러운 지수함수적 곡선으로 해석하느냐 아니면 두 개의 선형 관계와 혼합의 결과로 해석하느냐의 관점은 수괴의 해석에서 큰 견해차를 반영한다. 전자는 동해의 심해수를 동해고유수(Japan Sea Proper Water) 하나로 보는 일본 학자들의 견해를 지지하고 후자는 중층수 생성에 따라 다수의 심층수 수괴가 존재한다는 한국 학자들의 주장을 지지한다. Kido and Nishimura(1973)가 동해고유수(Japan Sea Proper Water)라 지목한 δt > 27.35인 심해수(> 800 m)에서 용존 규소의 농도가 증가하는 것이 뚜렷하게 보이는데 이를 하나의 수괴로 보는 것은 무리한 해석이라 판단된다. 1980년 후반부터 도입된 CTD-rosette 연속관측의 결과로 종래의 동해고유수 안에 비록 미약하지만 수온과 염분과 같은 보존적 속성의 연직분포에 확실한 차이가 있으며 비보존적이기는 하나 용존산소의 시간에 따른 변동성이 뚜렷하게 나타나는 것이 확인된 만큼 다수의 심층수가 존재하는 것에 무게가 실린다.
이어지는 토의에서는 수괴의 특성을 파악하는데 용존 규소의 연직분포가 시사하는 추가적인 정보와 일차생산에 대한 생지화학적 함의를 논의하였다.
4. 토 의
4.1 용존 규소의 농도와 동해 컨베어 벨트
동해 해수의 순환이 대서양의 남북연직대순환(AMOC)을 닮았다면 컨베어 벨트를 따라 영양염의 농도가 증가해야 할 것이라 예상할 수 있다. 그렇다면 울릉분지와 야마토분지 바닥에 가까운 > 2000 m 수심에서 동수심의 일본분지 보다 용존 규소가 높은 값을 보일 것이라 예상할 수 있다. 실제로 2001년에 연안에서 shelf cascading으로 저층수가 생성되는 것이 관측되었던 일본분지 저층수(>2300 m)의 인산염과 용존 규소의 농도는 울릉분지와 야마토 분지의 바닥에 해당하는 약 2300 m 수심의 값보다 같거나 약간 작게 나타나는 것으로 보이는데 차이가 뚜렷하지는 않다. 이러한 분포 양상에서 동해의 컨베어 벨트는 AMOC에 비해 분지 규모를 감안하더라도 규모가 훨씬 작음을 유추할 수 있다. 북대서양에서 침강한 북대서양 심층수(NADW)가 남극 대륙 가까이로 이동하는데 걸리는 시간 규모는 약 500년으로 추정되고 있다. 그런데 대서양 보다 규모가 훨씬 작은 동해의 컨베어 벨트의 순환 시간을 약 100년으로 추정하고 있다는 점은 심층수의 흐름이 상대적으로 느리며 이는 대륙붕수 침강(shelf water cascading)에 의해서만 저층수가 생성된다고 가정할 때 그 세력이 작음을 시사한다.
1980년대 후반부터 지금까지 동해를 관측한 자료는 해수 교체에 걸리는 시간 규모가 약 100년이라 할 때 이에 상응해서 심층수 체적의 1/100 규모로 매년 심층수가 만들어지는 것을 지지하지 않는다. 일본분지의 저층에서 용존산소가 꾸준히 감소하고 있는 점과 저층 혼합층(용존산소의 농도가 일정한 층)의 두께가 감소하고 있는 점에 비추어(Gamo et al., 2014) 저층수의 생성이 위축되고 있는 것은 확실하다. 한편 저층수 생성이 위축되는 것은 저층수 용승이 약화되는 것을 수반한다. 이는 플랑크톤의 내려보내기 생산(export production)이 표층으로 되돌려지기 어렵거나 최소한 시간 지연이 점차 길어질 것임을 시사한다.
그럼에도 동해 순환이 100년 규모로 계산되는 이유는 저층수 위에 존재하는 심층수의 생성이 활발하기 때문이라 해석되어야 한다. 심층수는 대륙붕수 침강 말고도 외양 대류(open ocean convection)로 만들어질 수 있기 때문이다. Yoon et al.(2018)은 온난화가 진행 중인 2000년대 이후에도 동해에서 심층수가 활발하게 만들어지는 증거를 제시한 바 있다. 여하튼 심층수의 생산은 일본분지 저층수에 들어있는 영양염이 표층으로 되돌려지는데 걸림돌이 될 것이 분명하다.
4.2 일본분지 저층에서의 일정한 용존 규소 연직분포
수심이 타 분지보다 깊은 일본분지의 심부(>2300 m)에서 용존 규소의 분포는 울릉분지나 야마토분지와는 달리 용존 규소의 농도가 눈에 띄게 수심에 따라 증가하지 않는다(Fig. 1(a) and (d)). 일반적으로 퇴적 표층에서 되돌려지는 용존 규소의 플럭스가 존재하는 점에 비추어 특이한 햔상이다. 우선 이 층의 두께는 정점에 따라서는 1000 m가 넘는데 가장 넓은 일본분지에서 장소에 따른 차이를 보이지 않는 점은 저층수 생성에 따른 비해 훨씬 우세한 수평적 혼합 메커니즘이 존재함을 시사한다. 조석에 따른 잔차류가 저층수 수평 혼합을 일으킬 가능성도 그 중 하나이다(Yang-Ki Cho, personal communication).
조석에 의한 혼합은 일본분지말고도 영향을 줄 것으로 예상되는데 실제로는 그렇지 않은 점은 해저지형과 수심의 영향으로 여겨진다. 일본분지의 지형은 비교적 단조로운데 반해 울릉분지와 야마토분지는 해산과 융기 대지가 존재해서 해저지형이 복잡하므로 저층 해류의 흐름이 제약을 받기 쉽다. 또한 수심이 얕은 두 분지로 유입되는 수괴는 저층수가 아니라 심층수여서 밀도 성층이 더 뚜렷하게 이루어져 있고 이것과 약한 저층수 움직임에 따른 혼합이 일본분지와는 달리 전 수심에서 용존 규소가 증가하는 양상을 빚어내는 것으로 여겨진다. 추가적으로 두 분지 저층에서 혼합층이 강하게 발달하지 않은 이유의 하나로 일본분지 저층수의 분지 유입이 없거나 미약하기 때문이라 해석할 수 있다.
일본분지 저층의 용존 규소 농도는 북대서양보다는 높고 북태평양보다는 낮다. 이는 AMOC 보다 세력이 약한 컨베어 벨트의 존재와 부합한다. 만약에 아표층수의 교체가 매우 활발하다면 용존 규소의 농도는 오히려 북대서양보다도 낮아야 할 것이다. 영양염이 고갈된 표층수가 심층으로 대량으로 수송되어 혼합될 것이기 때문이다. 일본분지 저층 용존 규소 농도의 최대값은 퇴적물에서 확산이 일어나는 바닥이 아니라 그 위에 존재한다. 이는 대륙붕수 침강으로 공급된 냉각된 표층수의 낮은 용존 규소 농도가 원인을 제공한 것으로 보인다.
4.3 일차생산에 대한 용존규소 제한 가설
용존 규소의 연직분포가 보이는 위로 볼록한 모습은 표면 혼합층 아래의 용존 규소 농도가 높은 아표층수가 용승 등의 과정을 통해 표면혼합층으로 원활하게 공급될 것이라 예상하게 하지만 인산염 대 용존 규소 비의 그림에서 보이는 추세선 기울기의 급격한 전환은 심층과 저층 영양염이 표층으로 공급되는 것이 강력한 성층에 의해 가로막힘을 시사한다. 밀도에 따른 용존 규소의 연직분포는 이런 예상을 뒷받침한다(Fig. 2). 용존 규소의 농도는 σθ ∼ 27.3부터 급격히 증가하며 이보다 가벼운 해수에서 농도는 거의 일정하게 20 이하로 낮다. 겨울철 외양 대류의 영향도 20μmol kg-1 이상으로 높여주지 않으며 혼합 수심도 500 m 보다 얕다. 특히 성층이 발달하는 고수온기에 심층수의 용존 규소를 비롯한 영양염이 표층에 공급되려면 상당한 밀도 차를 극복해야 하는데 국지적으로는 용승이나 소용돌이 혼합으로 가능할지라도 동해의 전반적인 분포 상황은 그렇지 않은 상태임을 볼 수 있다.
동해 심층의 영양염이 유광층으로 공급되기 어려운 상황이고 강에 의한 공급이 미미하므로 표면혼합층의 일차생산계는 표층 혼합층에서의 재생 플럭스와 대기 플럭스에 주로 의존해야 한다는 가정을 세울 수 있다. 그런데 용존 규소의 경우에는 추가로 대기 유입을 무시할 수 있다. 먼지에 포함된 규소 성분은 생규소와는 달리 풍화를 잘 견디기 때문이다. 따라서 일본분지에서 규조의 일차생산 기여도는 낮을 것으로 예상된다.
동해의 울릉분지와 일본분지의 상황은 조금 달라서 울릉분지에는 대마난류에 의한 공급이 있고 냉온수성 에디의 존재와 여름철 용승은 저층 용존 규소를 일부 이용할 수 있게 도와준다(Lee, 2018). 이에 반해 일본분지에는 외양 소용돌이가 잘 발달하지 않은데다 영양염이 소진된 표층 해수가 홋카이도 부근에서 아극전선 북쪽으로 진입하기 때문에 겨울철 혼합을 제외한 공급로가 없다고 보아도 무방해 보인다. 이러한 가정이 적절하다면 일본분지의 기초생산계는 용존 규소 수급에 어려움을 겪게 된다. 전 지구 규소 수지에 따르면 생규소의 약 1/4이 내려보내기 생산에 의해 저층으로 유입되고 그 가운데 10%가 넘게 표층퇴적물에서 용해되어 저층수로 돌아나오는데 제시한 가설에 따라 이 용존 규소가 표층으로 돌아오지 않고 저층에 갇히게 되면 용존 규소 수급에 대한 어려움이 시간이 갈수록 가중되게 된다. 가설에 따르면 일본분지의 기초생산에서 규조류의 기여도는 상대적으로 낮아야 하는데 아직 이를 지지할만한 기존 보고는 없다.
울릉분지의 경우에는 성층 시기에도 여름철 용승과 소용돌이 혼합이 대마난류의 공급과 더불어 표층으로의 용존 규소 공급을 도와줄 수 있을 것으로 예상된다. 그렇다 하더라도 울릉분지의 아표층에 가장 넓게 자리잡고 있는 모드 해수는 이러한 직관적인 해석을 복잡하게 만든다. 용승은 연안에서 좁은 띠로 단기간에만 일어나고 냉수성 소용돌이에 의한 에크만 펌핑의 혜택을 받는 면적도 그리 넓지 않다. 게다가 울릉분지에 존재하는 것으로 알려진 수온약층내 소용돌이(intrathermocline eddy)는 계절약층은 표층으로 부상시키는 대신 영구약층을 더 깊게 만들기 때문에 앞에서 언급한 표층과 심층의 격리를 강화시키는 역할을 하게 된다. 따라서 수온약층내 소용돌이가 해마다 모드수가 만들어질 때 영양염을 얼마나 가지게 되느냐가 관건이 된다. 이듬해 겨울에 모드수 형성시까지 기초생산을 지지하기에 충분한 양이라면 기초생산을 부양하게 될 것이고 반대라면 제한하게 될 것이다.
울릉분지의 아표층수에는 저온저염 해수 띠의 존재라는 변수가 하나 더 존재한다(Fig. 2 of Lee et al., 2012). 북한한류 또는 그로부터 파생된 해수로 보이는데 여름철에 고온저염의 표층수와 고온고염의 대마난류수 사이에 끼어든다. 두께가 얇은 층이지만 성층 시기에 유광대로의 저층 영양염 공급을 차단하는 효과를 낼 것이므로 기초생산에 심각한 제한을 가할 것으로 예상된다 메타게놈 분석에 따르면 이 시기에 울릉분지에서 일차생산자의 종조성은 와편모조가 주도하고 규조의 다양성은 매우 빈약하다(Fig. 3 of Lee et al., 2012). 메타게놈 분석은 정성적인 결과라서 정량적으로 확대해석하는 데에는 주의가 필요하지만 규조의 종 다양성이 빈약한 것은 일단 용존 규소의 공급이 원활하지 못 할 것이라는 가정과 맥락을 같이한다.
결론적으로 동해는 분지 별로 원인에는 차이가 있으나 현재 심해로 송출된 용존 규소가 표층으로 되돌아오기 어려운 여건 아래에 있으며 지구온난화에 따라 상황이 더 심각해질지는 중요한 연구 대상이라 판단된다.
4.4 동해에서 용존 규소의 생지화학적 순환이 지닌 함의
가장 최근에 규소 순환 연구를 종합 검토해서 내린 결론 가운데 하나는 전 지구 해양 순환에 있어서 규소는 정상상태에 있지 않으며 체류시간도 과거의 추정보다 짧은 10,000년 규모이고 기후변화와 같은 인위적인 외압에 민감하며 규소의 재생은 심해에서 침강입자의 분해가 아닌 표층 해양과 해수-퇴적물 경계면에서 주로 일어난다는 것이다(Treguer and De La Rocha, 2013).
대지이용의 변화와 같은 인간의 외압, 특히 댐의 건설은 해양으로의 용존 규소 공급을 줄여왔기 때문에 연근해에서 가장 뚜렷하게 나타나며 향후 예상되는 지구온난화에 대해 민감하게 반응을 보일 것이다. 동해의 경우에는 육상 공급이 줄어든 것에 더해 보다 중요한 공급원으로 지목되고 있는 심해 저장소에서 유광대로의 유입 플럭스가 지구온난화 등 기후변화에 의해 해수 교체 양상이 불리한 쪽으로 바뀌고 있는 것으로 보여서 용존 규소 제한이 심화될 것이라는 예상을 하게 된다.
규조의 외각이 두꺼울수록 풍화를 잘 견디므로 탄소를 심해로 전달하는 효율이 높아지게 된다. 하지만 규조의 성장이 빨라서 얇은 외각을 만든 경우에는 초식자의 섭이와 분립을 통해 송출하게 되고, 해설로 응집되어 빠르게 침강할 확률이 높아지므로 역시 생물학적 탄소 펌프가 강화된다. 결과적으로 규조의 존재는 성장 속도와 무관하게(외각의 두께와 관계없이) 각기 다른 메커니즘을 통해 탄소의 심해 송출을 촉진한다. 그러므로 용존 규소 제한은 규조의 생체량 감소로 이어져서 궁극적으로는 생물학적 탄소펌프를 약화시킬 것이라 예상할 수 있다. 생태계가 이에 반응해서 미생물 순환 고리를 강화시켜 생물학적 탄소펌프를 유지시켜 줄지도 모르지만 결과를 전망하기에는 불확실성이 여럿 존재한다. 만약에 기후변화의 영향으로 하위영양단계에서 생규소나 탄산칼슘 껍데기를 가진 플랑크톤의 비중이 준다면 생물학적 탄소펌프에 대한 입자꼴 유기탄소(POC)의 기여는 줄고 용존태 유기탄소(DOC)가 늘어나게 될 것인가 하는 질문이 제기된다. Kim et al.(2017)은 동해 해수의 유기탄소 농도가 대양에 비해 월등하게 높음을 발표한 바 있고, Chen et al.(2017)은 온난화로 해수 교체가 느려지더라도 침강한 유기물의 효율적인 분해를 통해 심해수의 산성화가 촉진될 것이라고 동해를 예를 들어 제안하였다. 이러한 맥락에서 온난화된 해양에서 내려보내기 생산이 광물질 부력조절(ballasting) 주도에서 껍데기가 없는 미소생물의 점착물(aggregatea)로 바뀌고 해양의 생지화학적 순환에서 저층수의 기여가 저감되는 방향으로 진행될지 여부는 커다란 관심사로 떠오르게 될 것이라 전망할 수 있다. 단 Baines et al.(2010)이 보고한 바와 같이 규소를 함유한 초미세플랑크톤이 생규소 생산의 60% 가량을 차지한다면 생규소의 순환은 지금까지 규조가 주도한다는 패러다임이 수정되어야 하므로 주의가 필요하다.
이 논문에서는 세포의 원형질을 이루는 인산염과 껍데기를 만드는데 쓰이는 용존 규소 사이의 비가 보이는 연직분포의 변화를 분해(용해)와 심해수 형성 메커니즘과 연관지어 설명하고자 하였다. 그 결과 현재 동해 심층수 형성 모드는 내려보내진 규소가 표면 혼합층으로 원활하게 되돌리기 어렵게 만든다는 결론에 도달했다. 동해의 생물펌프가 입자꼴 유기물 침강 주도에서 용존 유기물 위주로 바뀔지도 모른다는 가설을 제기하게 되었는데 미래의 온난화된 해양이 동해의 전철을 밟게 될는지는 지대한 관심사이다. 한편 기존의 규조 중심의 규소 순환 패러다임이 최근에 도전을 받고 있는 바 향후 동해에서 규소 순환의 주도적인 생물군에 대한 연구가 추가적으로 필요하다고 생각된다.
