

1. 서 론
2. 재료 및 방법
3. 결과
3.1 해역별, 수심별 연체동물의 섭식형 조성율(%)
3.2 해역별, 수심별 해조류 군락 특성
3.3 해역별 투명도
3.4 해역별 식물플랑크톤 밀도
4. 고 찰
1. 서 론
암반(경성기질) 조하대에 서식하는 저서동물을 포함한 모든 동물의 생존에는 먹이가 필수적인 요소인 만큼 섭식 가능한 먹이에 대한 강한 선택성과 섭식의 효율성(optimal foraging)은 기본적 필요 사항이며, 거시적 관점에서 섭식 가능한 먹이의 종류와 양이 암반 조하대 저서동물 생존에 필수적인 요소로 작용함은 분명하다. 그러므로 암반 생태계는 생산자로서의 먹이 종류와 양에 따라 소비자로서의 저서동물 종류, 밀도 등과 유의한 상관성을 갖는 역학적 구조를 나타내게 되며(Kwon et al., 2010), 그 곳에서의 적합한 섭식형과 해당 섭식형을 가진 종들이 우점하는(‘guild structure’ in Root, 1967) 군집구조가 형성된다.
암반 조하대에 서식하는 대부분의 저서동물들은 영구부착성이거나 이동성이 낮아서 연안역의 환경변화를 추적하는 주요한 대상 생물군으로 평가받지만(Wenner, 1987; Lindeman et al., 2009; Dauvin et al., 2010), 직접적인 잠수작업과 수중에서의 정량적 표본채집의 어려움 등(Bohnsack, 1979)으로 인하여 연구의 빈도와 강도는 연성기질 조하대에 서식하는 저서동물 연구(Yoon et al., 2009; Maeng et al., 2015; Lim et al., 2018)에 비해 상대적으로 매우 낮은 편이다. 제한적이고 까다로운 연구 조건에도 불구하고 조하대 경성기질 저서동물 군집은 주로 서식하는 수심의 차이에 따라 서로 다른 구조적 특징을 나타내는 것으로 알려져 있다(Ruitton et al., 2000 in Patella spp. community). 일차적으로 바닷속으로 유입되는 수심별 광량의 차이는 수심에 따른 해조류의 생체량과 군락의 구조적(flora) 차이를 가져오게 되고(Grace, 1983), 나아가 이들 해조류를 섭식하는 초식형 저서동물의 우점 또는 비우점이라는 군집구조의 차이를 유발한다(Ruitton et al., 2000). 또한 인과관계를 명확히 제시하지는 못했으나 수심이 깊어질수록 부유물 여과섭식형 저서동물인 태형동물의 우점정도가 강해진다는 연구결과도 제시된 바 있다(Grace, 1983).
한국 연안 암반 조하대 저서동물 군집구조에 대한 연구는 Son et al.(2004a)과 Kwon et al.(2010)등과 같은 일부 연구를 제외하고는 지금까지 군집구조 그 자체에 대한 기술적(descriptive) 연구(Seo et al., 2009; Yoon et al., 2009; Lim et al., 2018) 또는 서식 종들에 대한 동물상 목록(inventory)을 파악하는 등에 대한 연구들(Son et al., 2004b; Choi et al., 2006)이 대부분을 차지하고 있다. 그러나 암반 조하대 저서동물 군집구조를 포함한 “대부분의 군집구조는 군집 내에서 나타나는 그들 고유의 먹이망에 의해 결정되고 유지된다.”(Pianka, 1982)는 이론을 생각할 때, 반도의 절반 이상을 차지하는 한국 연안의 암반 조하대 저서동물 군집구조와 그 결정 요소에 대한 실증적 연구도 필요하다는 것은 명백한 사실이다.
따라서, 본 연구의 목적은 먹이망에 기초하여 한국 연안 암반 조하대 저서동물 중 연체동물을 대상으로 먹이생물의 종류와 양이 연체동물 군집구조를 결정하는 하나의 요인이 될 수 있는지를 확인하는 데 있다.
2. 재료 및 방법
암반 조하대 저서동물 및 해조류 표본은 한국 연안에 유사한 수평간격으로 설정된 총 18개 정점(Table 1, Fig. 1)에서 채집되었다. 각 정점에서의 표본 채집은 저서동물 및 해조류의 신규 개체군 가입정도와 성장 및 생장 시기 등의 차이에 따른 군집 또는 군락의 구조적 변이를 최소화하기 위하여 연중 동일한 시기인 5월과 8월(2015년과 2016년 또는 2017년), 총 2회에 걸쳐 수행되었으며, 총 2회의 결과 자료들은 모두 통합되어(data pooling) 분석되었다. 또한, 각 정점별 수심 차이에 의한 군집구조의 현상왜곡을 방지하기 위하여 표본을 각 정점의 수심 5 m(- 5 m)와 15 m(- 15 m)에서 매회 2번씩 정량적으로 반복 채집, 분석하였다. 따라서, 각 해역별 총 채집면적은 동해 14 m2, 남해 10 m2, 황해 12 m2로 차이를 보이며 이에 따라 출현 종 수 차이의 변이를 고려하여 실제 분석에서는 총 출현 종 수에 대한 각 섭식형의 조성율(%)을 기준으로 비교하였으며 각 해역별, 수심별 총 출현 종 수는 참고로 제시하였다. 특히, 동해에 비해 상대적으로 조차가 큰 남해와 황해의 경우, 수심 차이에 따른 현상왜곡을 방지하기 위하여 매 조사 시점을 5월의 소조기 정조 시점으로 통일하여 표본 채집을 수행하였다. 표본 채집 정점(들) 위치의 해역별 구분은 Park(1970)의 한국 연안 해역 구분을 따랐다.
Table 1. Sampling site, in detail, of hard-bottom subtidal mollusks and algae in the Korean coasts on May and August from 2015 to 2017 with a specified underwater quadrate and scuba diving
각 정점과 수심에서의 정량적 표본채집은 scuba diving을 통해 표본 채집 시 1-2 mm 크기의 소형 표본개체들의 유실방지를 위하여 특별히 고안된(Son et al., 2004a) 수중방형구와 끌칼을 이용하여 방형구 접지면적(50 × 50 cm) 내 존재하는 모든 생물체(크기 1 mm 이상의 대형 저서동물 및 해조류)를 채집하는 파괴적 방법으로 이루어졌다. 접지면적 내 존재하는 모든 표본들은 방형구에 직접 연결된 망목크기 1 mm의 표본채집망에 모은 후 수중에서의 이동과정에서 발생 가능한 표본유실 방지를 위하여 채집망을 완전히 밀봉한 상태로 선상으로 운반하였으며, 운반된 표본은 미리 얼음을 채워둔 아이스박스에 보관하여 실험실로 운반하였다. 실험실로 운반된 저서동물 표본들은 일차적으로 패각 부스러기나 자갈과 같은 비생물을 선별, 제거한 후 적정크기의 고정 및 보존용기에 넣고 Borax로 중화시킨 10% 해수-포르말린 수용액으로 고정, 보존하였다. 해조류의 경우, 앞의 비생물 선별 과정에서 별도로 해조류만을 선별한 후 저서동물과는 별도의 적정용기에 넣은 후 종 동정 및 생체량 계측 시까지 냉장 보관하였다.
최소 7일 이상 고정-보존된 저서동물 표본은 담수로 충분히 세척한 후 1차 대분류과정에서 연체동물만을 선별하여 각 종을 동정하였다. 동정된 종에 대해서는 다양한 참고문헌(예. Barnes and Hughes, 1988; Shimeta and Jumars, 1991)과 실제 실험실 내 사육 경험 등에 근거하여 각 종별 섭식형을 결정하였으며, 단일 섭식형이 아닌 종의 경우(예. 얼룩덜룩갈줄고둥, Cerithium alutaceum: 퇴적물섭식형이자 저서규조류도 섭식하는 초식형) 해당 종의 주된 섭식형(얼룩덜룩갈줄고둥: 퇴적물섭식형)으로 분류하여 분석하였다. 냉장상태로 보관된 해조류 표본의 경우 종 동정과 함께 각 개체별 생체량(건중량)을 전자저울(CAS-MWP, Korea)을 이용하여 소수점 2자리까지 측정하였으며, 분석된 자료들을 대상으로 Litter et al.(1983)의 기준에 따라 6개의 기능형으로 구분하여 최종 분석에 이용하였다.
투명도는 ‘국가해양환경측정망조사’(2015-2017)(http://www.meis.go.kr/rest/main) 자료를 바탕으로 암반 조하대 조사정점과 가장 인접한 조사정점에서 5월과 8월에 측정된 최근 3년간의 자료를 평균으로 산출하여 최종 분석 및 결과에 대한 고찰에 사용하였다(Table 2).
Table 2. Survey stations of the transparency in the Korean coasts. Data were collected from MEIS data bank (2015-2017; http://www.meis.go.kr/rest/main)
| Location | Station |
| East Sea | Yangyang-01, Hupo-02, Jukbyeon-06, Busan-09, Gampo-02 |
| South Sea | Deungnyang-05, Namhaedonaman-04, Geojedodongan-03, Wando-04 |
| Yellow Sea | Jeonjupo-03, Daesan-04, Boryeong-02 |
본 연구의 대상인 암반 저서동물들은 그들의 섭식형 중에서 여과섭식형의 경우, 촉수의 수관이나 미세 곁가지를 이용하여 여과섭식하는 극히 일부 종들(예. 일부 거미불가사리, 말미잘 등)을 제외한 대부분의 종들이 식물플랑크톤을 먹이로 한다(Meyer, 1973). 따라서, 본 연구의 대상인 여과섭식형 연체동물의 대부분은 식물플랑크톤을 먹이로 한다는 사실에 기초하여 해조류 외, 식물플랑크톤만을 분석 대상으로 하였다. 식물플랑크톤 밀도는 ‘국가해양생태계종합조사(연안생태계)’(2015-2017) (http://www.meis.go.kr/rest/main) 정점 중, 암반 조하대 조사정점과 가장 인접한 조사정점에서 최근 3년간 5월과 8월에 채집, 분석된 결과들을 평균으로 산출하여 최종 분석 및 결과에 대한 고찰에 사용하였다(Table 3).
Table 3. Sampling stations of the phytoplankton in the Korean coasts. Data were collected from MEIS data bank (2015-2017; http://www.meis.go.kr/rest/main)
3. 결과
3.1 해역별, 수심별 연체동물의 섭식형 조성율(%)
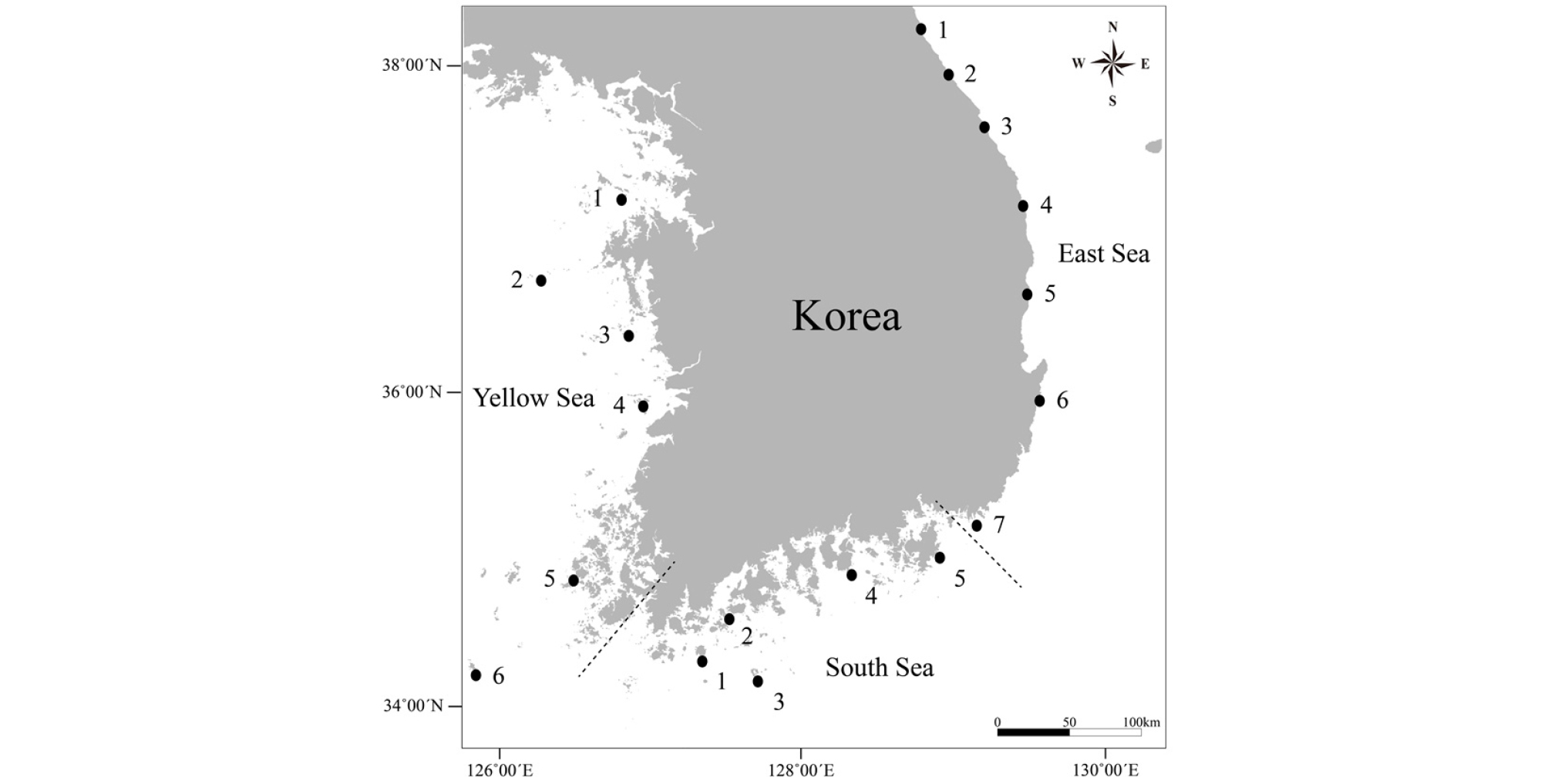
동해의 경우, 암반 조하대 상부(이하 ‘상부’)에서 총 75종의 연체동물이, 그리고 암반 조하대 하부(이하 ‘하부’)에서 총 105종의 연체동물이 출현하였다. 이들 중 가장 우점적인 섭식형은 초식형으로 상부에서 52.0%(39종), 하부에서 43.8%(46종)를 차지하였다. 상부와 하부 모두에서 최우점한 초식형 다음으로 포식형→여과섭식형→퇴적물섭식형의 순으로 나타나 섭식형의 상대적 조성율(%)의 순위에 있어서 상부와 하부의 차이가 없는 것으로 나타났다(Table 4, Fig. 2). 최우점 섭식형인 초식형 조성율의 상․하부 산술적 평균값은 차우점 섭식형인 포식형의 약 2배에 해당되어 동해 암반 조하대 서식 연체동물의 경우 초식자들이 절대적으로 우점한 군집구조를 나타내었다(Table 4).
Table 4. Spacial and vertical variation of percent composition (%) of the mollusk's feeding type based on the quantitative hard-bottom subtidal samples for 2015-2017 with scuba diving and a specified underwater quadrate in the Korean coasts
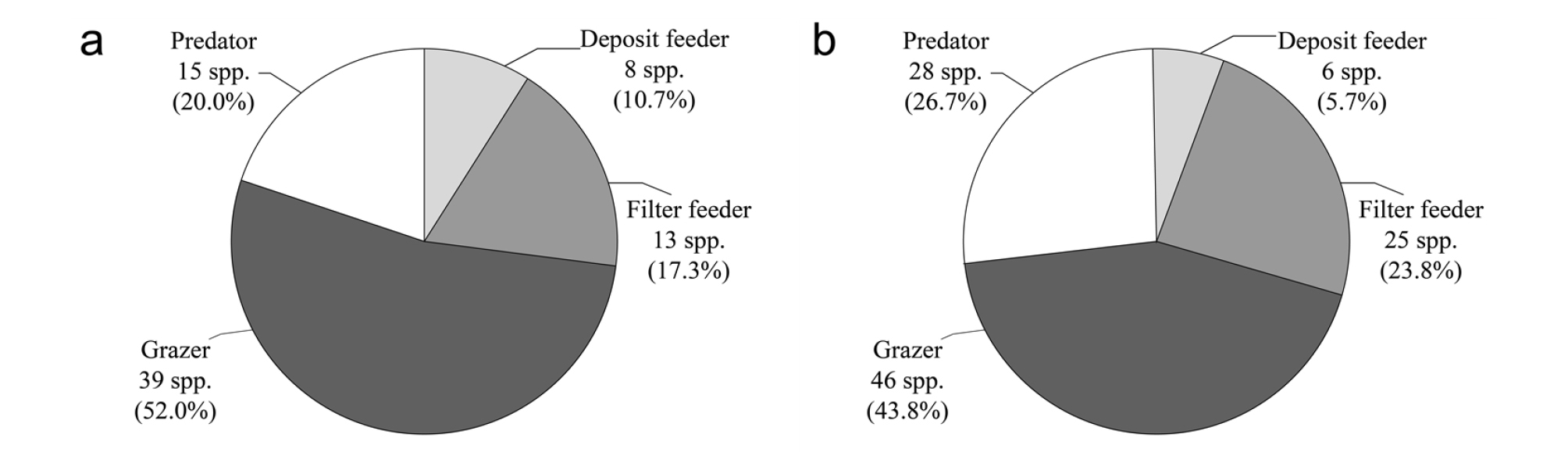
남해의 경우, 상부에서 총 82종, 하부에서 총 74종의 연체동물이 출현하였다. 이들 중, 상부에서 가장 우점한 섭식형은 초식형으로 34.2%(28종)를 차지하였고 다음으로 여과섭식형(28.0%, 23종)→포식형(25.6%, 21종)→퇴적물섭식형(11.0%, 9종)→잡식형(1.2%, 1종)의 순서로 나타났다. 반면 하부에서는 상부와는 달리 포식형(33.8%, 25종)이 가장 우점적인 섭식형으로 나타났으며, 다음으로 여과섭식형(31.1%, 23종)과 초식형(31.1%, 23종)이 동일한 조성율을 나타내었고, 퇴적물섭식형이 가장 낮은 조성율(4.0%, 3종)을 나타내었다(Table 4, Fig. 3). 따라서 남해의 경우 동해와는 달리 최우점 섭식형인 초식형의 차지 비율(상․하부 비율의 산술적 평균값)이 32.6%로 여과섭식형(29.6%) 및 포식형(29.7%)과 거의 유사한 값을 나타내어 어느 특정 섭식형이 절대적 우점을 나타내는 군집구조가 아님을 알 수 있었다(Table 4).
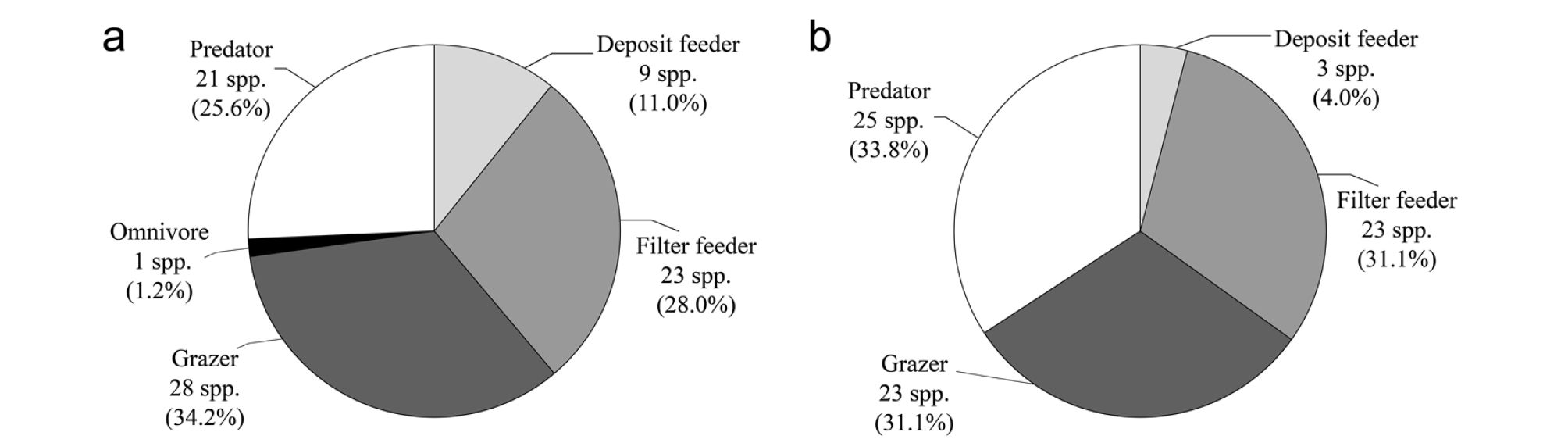
황해의 경우, 상부에서 총 76종, 하부에서 총 54종의 연체동물이 출현하였다. 이들 중, 가장 우점한 섭식형은 여과섭식형으로 상부에서 38.2%(29종), 하부에서 46.3%(25종)를 차지하였다. 섭식형에 있어서 황해의 경우 상부와 하부에서 서로 상이한 조성율을 나타내어 최우점적인 여과섭식형 다음으로 상부에서는 초식형→포식형→퇴적물섭식형으로 나타난 반면, 하부에서는 포식형→초식형→퇴적물섭식형의 순서로 나타났다(Table 4, Fig. 4). 최우점 섭식형인 여과섭식형 상․하부 비율의 산술적 평균값이 차우점 섭식형인 초식형(25.7%) 및 포식형(23.2%)에 비해 약 2배 가량 높아 여과섭식자들이 절대적으로 우점하는 군집구조를 나타내고 있었다(Table 4).
3.2 해역별, 수심별 해조류 군락 특성
해역별 해조류 생체량은 동해와 황해에 비해 남해에서 가장 많았으며, 수심별로도 남해에서는 특정 수심에 집중적으로 생체량이 많은 것이 아니라 상․하부 모두에서 생체량이 많았다(Table 5). 황해 하부의 해조류 생체량은 동․남해 하부 대비 10% 미만의 생체량을 보였으며 해역별 및 수심별 해조류 생체량 중 가장 적었다.
Table 5. Spacial and vertical variation of algae biomass with total and two functional forms (sheet (S) and thick-leathery (TL) forms; Litter et al., 1983). Stations represent sum of the number of sampling stations in the each location. Data of the each location were pooled for analysis. Mean represents means of biomass (gDW/m2) of all algae and of the two functional forms at the two vertical positions (- 5 and - 15 m), respectively
한편 전체 출현 해조류 중 초식형 연체동물의 주된 먹이자원이 되는 엽상형과 다육질형 종들이 차지하는 비율에 있어서는 동해가 남해와 황해 대비 각각 약 1.3배와 2.1배 높은 값을 나타냄으로써 동해 암반 조하대에 서식하는 해조류는 다른 해역에 비해 주로 엽상형과 다육질형 종들이 차지하고 있음을 알 수 있었다(Table 5).
3.3 해역별 투명도
해역별 투명도는 동해(7.1±3.0 m)→남해(4.0±2.1 m)→황해(3.2±1.6 m)의 순서로 높게 나타났다(Fig. 5). 특히, 동해에서는 황해 대비 2배 이상 높은 값을 나타내어 투명도가 확연히 높았다. 그러나 남해와 황해의 경우, 남해의 투명도가 상대적으로 높긴 하지만 그 차이가 0.8 m 정도에 불과하여 서로 큰 차이를 보이지 않았다.
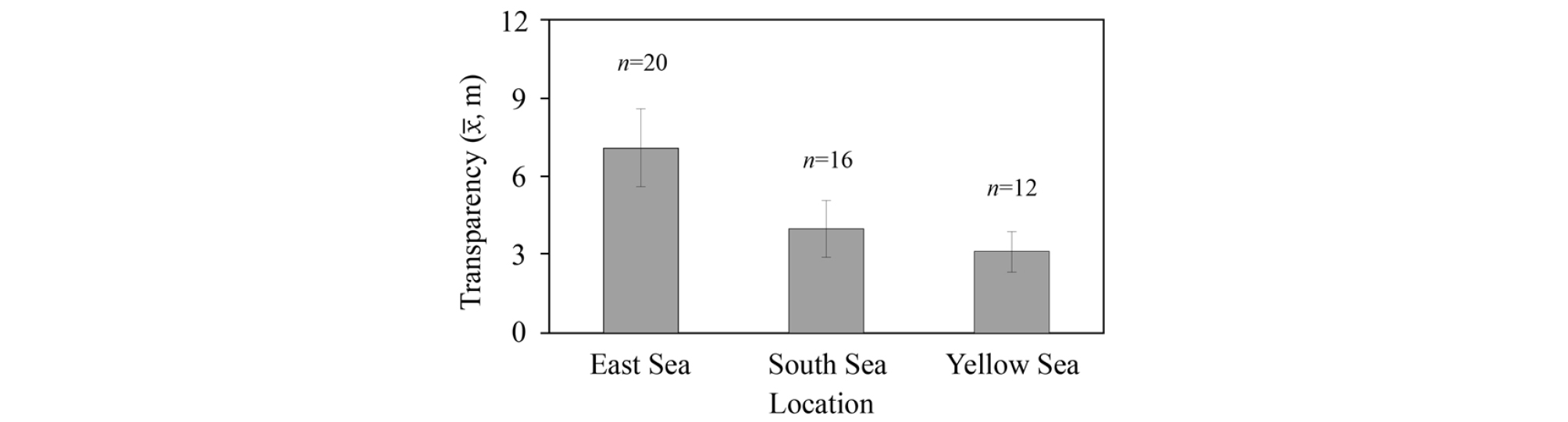
Fig. 5.
Spacial and vertical variation of algae biomass with total and two functional forms (sheet (S) and thick-leathery (TL) forms; Litter et al., 1983). Stations represent sum of the number of sampling stations in the each location. Data of the each location were pooled for analysis. Mean represents means of biomass (gDW/m2) of all algae and of the two functional forms at the two vertical positions (- 5 and - 15 m), respectively
3.4 해역별 식물플랑크톤 밀도
해역별 식물플랑크톤 밀도는 남해→동해→황해의 순서로 높게 나타났다. 그러나 동해와 남해의 식물플랑크톤 밀도는 큰 차이를 보이지 않았으며, 황해에서는 타 해역에 비해 상대적으로 낮은 밀도를 보였다(Table 6).
Table 6. Spacial variation of the phytoplankton density in the Korean coasts on May and August from 2015 to 2017. Location in detail see the Table 3
Location Variable | East Sea | South Sea | Yellow Sea |
| Density (mean, cells/mL) | 353±530 | 378±454 | 297±233 |
| Number of station | 7 | 5 | 6 |
4. 고 찰
자연에 서식하는 많은 종들은 특별한 한 종류의 섭식형을 갖기보다는 생존에 보다 유리한 복합적 섭식형을 가진다(Levinton, 1982). 그러나 복합적인 섭식형을 가진 종이라도, 일정 크기 이상의 먹이는 섭식 대상으로 선택하지 않는 등의 복잡한 섭식 기작(Brown, 1997)과 자신의 섭식 효율을 극대화(optimal foraging)시킬 수 있는 매우 정교한 먹이선택성(DeMott, 1989)을 가진다. 따라서, Levinton(1982)의 연구 등 기존 연구 결과들을 바탕으로 암반 조하대 연체동물의 각 종별 섭식형을 초식형, 여과섭식형 또는 퇴적물섭식형 등으로 저자들이 단정적으로 판단하여 먹이자원의 종류 및 양과 결부시켜 분석하고 해석하는 것에 무리가 있을 수 있다.
그러나 Root(1967) 이후 자연적인 현상으로 “동일 또는 유사한 먹이자원에 의해 동일한 섭식형의 종들로 구성되는”(Mcdonald et al., 2010) “Feeding guild”의 개념과 그 실체에 관한 연구(Kwon et al., 2010)가 있다. 특히, 한국의 경우 외해와 바로 연결되는 단순한 해안선을 가진 동해, 다수의 도서들에 의한 거시적 관점에서의 반폐쇄적인 남해 그리고 큰 조차를 가진 황해가 공존하는 자연환경을 보유하고 있어 다양한 해양환경의 조건에서 “Feeding guild”의 개념을 적용시킬 수 있었다.
“Feeding guild”의 개념에 기초한 암반 조하대 서식 연체동물 군집구조의 결정 요소로서 이용 가능한 먹이자원에 대해 살펴본 본 연구는 초식형의 먹이인 해조류와 여과섭식형의 먹이인 식물플랑크톤에 한정하여 분석한 한계가 있다. 그러나 이러한 한계성에도 불구하고 본 연구는 Feeding guild의 개념을 뚜렷하게 증명하고 있었다. 동해의 경우 초식형의 절대적 우점현상(상․하부 자료 통합, 47.9%, Fig. 6a)은 해조류 중 저서동물의 섭식 이용 빈도가 높은 엽상형과 다육질형(Yoo et al., 2007; Kim and Jang, 2012)이 약 40%를 차지하고 있다는 점에서(Table 5) 먹이자원에 의한 “Feeding guild”의 형성과 그에 따른 군집구조 결정의 상호 연관성을 보여준다. 남해와 거의 유사한 식물플랑크톤 밀도를 나타낸 동해에서 여과섭식형의 상대적 비율이 초식형의 약 1/2 밖에 되지 않는(Fig. 6a) 이유 중 하나는 한정된 이차원적 암반 생태계 내에서 초식자들의 공간적 선점 또는 우점이 여과섭식자들의 먹이자원이 충분히 존재함에도 불구하고 이들의 개체군 규모를 제한하는 요인으로 작용할 가능성도 있을 것으로 생각된다. 그러나 초식자의 섭식행동으로 인해 먹이자원인 다생 해조류 종이 초식자에 대한 방어물질 생산을 촉진한다는 연구 결과(Van Alstyne, 1988)를 고려할 때, 초식자들이 지속적으로 우점할 것인지에 대해서 보다 장기적인 연구도 필요할 것으로 생각된다. 남해의 경우 타 해역 대비 특정 먹이자원의 두드러진 우세 현상이 나타나지 않아 초식자, 포식자, 여과섭식자가 다양하게 공존하고 있었으며(Fig. 6b), 이는 먹이자원과 저서동물(연체동물) 군집구조와의 관련성을 논리적으로 보여주는 하나의 실증 사례라고 생각된다.
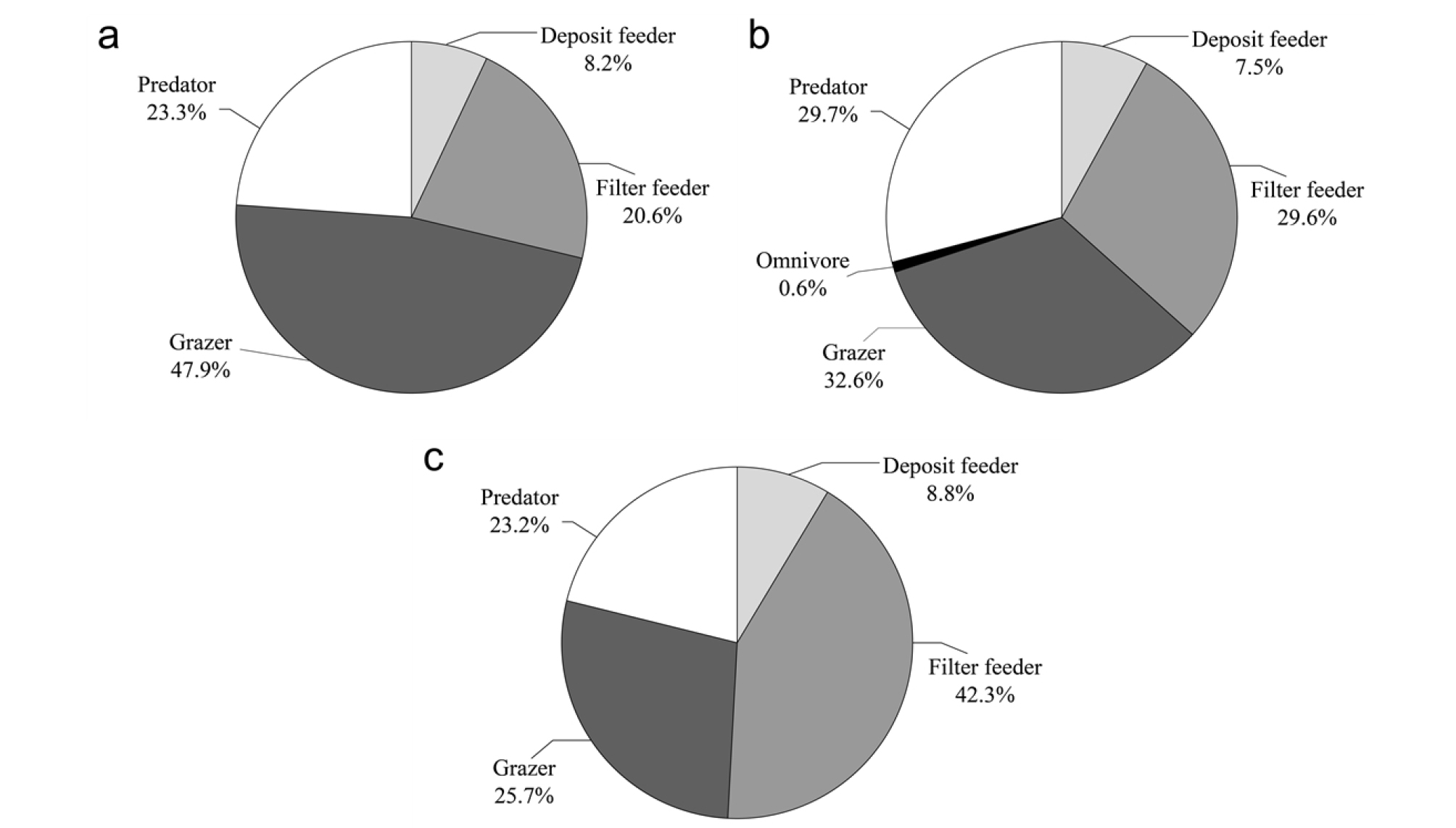
한편, 특정 섭식형의 우점현상이 타 섭식형 종들의 개체군 규모에 대한 제한 요인으로 작용할 수도 있다는 추론은 황해에서도 적용 가능하다. 황해의 경우, 해조류의 전체 생체량이 가장 낮았으며(Table 5), 그 결과로 추정되는 초식형 연체동물의 조성율 역시 세 해역 중 가장 낮은 25.7%를 나타내고 있었다. 하지만 식물플랑크톤의 상대적 밀도는 세 해역이 유사하여 여과섭식자들의 공간적 우점 또는 선점이 앞서 이루어짐으로써 초식자들의 개체군 규모를 제한한 것으로 생각된다(Table 6, Fig. 6c).
여과섭식형 종들 중 상당수는 먹이자원인 식물플랑크톤의 밀도와 서식 수온 등에 따라 여과섭식율이 달라질 수 있고(Okumus et al., 2002) 개체발생 과정에서 먹이의 크기를 변화시킨다(Burns, 1968). 이러한 사실은 본 연구가 온대 해역에서의 사계절 중 단지 5월(봄)과 8월(여름)에만 실시되어 계절적 변동 양상에 대한 정보가 제한됨으로써 여과섭식형 종들의 개체군 신규 가입과 성장, 생존 등을 명확히 파악하지 못한 한계를 가지고 있다. 또한 물리적 또는 기계적으로 동일한 여과기작에 의한 섭식 방식을 지닐지라도 섭식자의 종에 따라 또는 세부적인 여과기작의 차이에 따라 먹이 크기를 달리하며, 먹이자원인 식물플랑크톤도 동일한 상위 분류군(higher taxa)에 속하더라도 각 종의 크기는 매우 다양하다(Werner and Werner, 1954). 따라서 향후의 연구에서는 섭식형의 보다 세분화 및 식물플랑크톤의 크기 세분화(size fraction)를 바탕으로 상호 연관성에 대한 추가적 연구도 필요할 것으로 생각된다.
본 연구의 방법과 결과가 제한적이기는 하지만 “우리나라 암반 조하대 연체동물 군집구조의 결정에는 해당 서식지에서의 이용 가능한 먹이자원의 종류와 양이 하나의 요소로 작용하고 있다.”는 결론 도출이 가능할 것으로 판단된다.
